Ljubljana/model
From 2007.igem.org
Model
Our aim was to build a theoretical model to show how the feedback loop made of T7 RNAP gene under control of its T7 promoter affects the behaviour of the system, particularly amplification. Initially, active T7 RNAP molecules are generated by any of three sources: split-ubiquitin formation, TEV protease reconstitution or cleavage off the membrane by HIV protease. In all cases, T7 polymerase translocates into the nucleus, where it transcribes an effector gene, as well as T7 RNAP gene (self-amplification). The idea behind introducing such a positive feedback loop was that the initial signal might be too low and could fade if not amplied to a sufficient level.
Model in Cell Designer
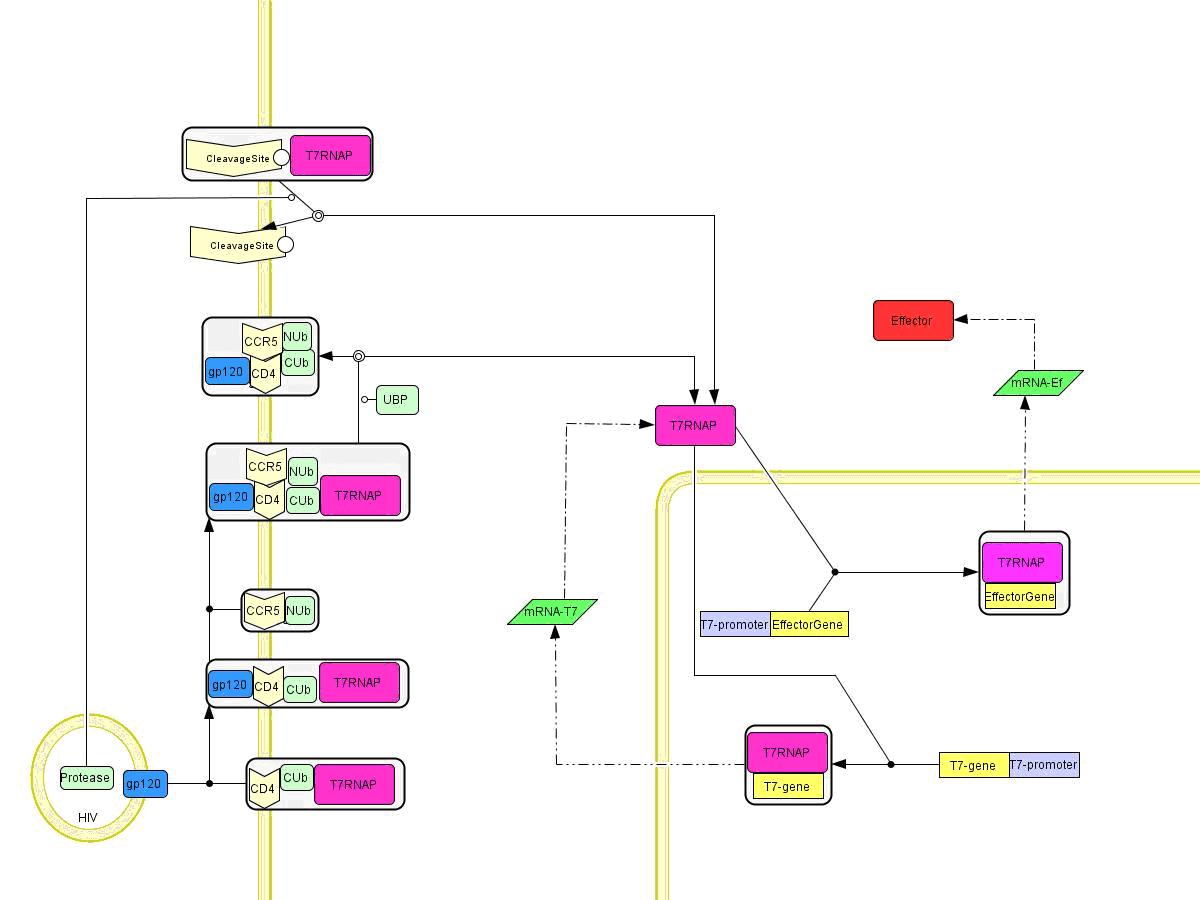
Figure 1: Comprehensive view of the engineered pathways. Split ubiquitin pathway is shown on the lower half and HIV protease on the upper half. HIV protease synthesis pathway after infection is simplified in this model.
Signal Amplification
Plots of amounts of active T7 RNAP (red line) and effector (blue line) versus time. The infection begins at time 0. Parameters are selected arbitrarily in order to present the general behavior of the system:
|
|
|
|
|
|
|
|
|





As expected, in case (a), where no amplification through T7 RNAP was designed, the amount of effector increases linearily with time. With more copies of the effector gene present, the slope increases (b). In case (c) with amplification through T7 RNAP, we were expecting the effector amount to grow exponentially. However, contrary to simple reasoning, this is not the case unless the number of effector gene copies is much larger than the number of T7 RNAP gene copies. This can be explained by a limiting rate of transcription; the maximal transcrition rate is determined by how many polymerase molecules can bind to the promoter in a certain span of time. Therefore, if the number of polymerase molecules is already high, the system becomes saturated very soon, and any further increase of polymerase concentration does not increase expression rate, so no exponential growth of effector concentration occurs. However, if we increase the number of effector gene copies (d), the system does not readily become saturated and we observe an exponential growth of effector concentration. After a certain time, the number of polymerase molecules increases, and effector genes become saturated. This again results in a linear growth of effector concentration.
The model clearly showed that in order to successfully amplify the initiatial signal, it is optimal to use a much larger number of effector genes than of T7 RNAP genes.